Research article
Association of the virulence factors of Helicobacter pylori and gastric mucosal interleukin-17/23 mRNA expression in dyspeptic patients
Nader Bagheri1, Ghorbanali Rahimian2, Loghman Salimzadeh1, Fatemeh Azadegan1, Mahmoud Rafieian-Kopaei3, Afshin Taghikhani2, Hedayatollah Shirzad1[*]
1Cellular and Molecular Research Center, Shahrekord University of Medical Sciences, Shahrekord, Iran2Dept. of Internal Medicine, Shahrekord University of Medical Sciences, Shahrekord, Iran
3Medical Plants Research Center, Shahrekord University of Medical Sciences, Shahrekord, Iran
EXCLI J 2013;12:Doc5
Abstract
The molecular pathways that control Helicobacter pylori (Hp)-associated inflammatory reaction are complex, but locally induced cytokines and virulence factors seem to have a major role in maintaining the ongoing inflammation. Therefore this study was aimed to evaluate the association of the virulence factors of Hp and gastric mucosal interleukin-17/23 mRNA expression in dyspeptic patients.
Mucosal IL-17 and IL-23 mRNA expression in H. pylori infected and non-infected gastric biopsies were determined by real-time RT-PCR. Virulence factors, vac-A and cag-A were evaluated using PCR.
There was no significant difference in mucosal IL-17 and IL-23 mRNA expression between H. pylori infected and non-infected patients. Their expression in mucosa did not correlate with chronic gastritis and chronic active gastritis. IL-17 and IL-23 mRNA expression in mucosa of patients with vacA m1 were significantly higher than those observed in patients with vacA m2. The severity of polymorphonuclear infiltration and chronic active gastritis was higher in cag-A positive than cag-A negative patients.
H. pylori infections carrying the vacA m1 allele have higher IL-17 and IL-23 mRNA and the current study suggests that the virulence factor vacA allele's m1 are important for the severe gastric inflammation.
Keywords: H. pylori, virulence factor, interleukin, gastritis
Introduction
H pylori is a spiral-shaped Gram-negative flagellate bacterium that colonizes the gastric mucosa of approximately 50 % of the world's population. Although its colonization often stays asymptomatic, but can progress into gastric or duodenal ulcers, gastric malignancies and mucosa-associated lymphoid tissue lymphomas (Fox and Wang, 2007[9]). Hp-associated inflammatory reaction is defined by a massive mucosal infiltration of polymorphonuclear leukocytes (PMN), T cells, macrophages, and plasma cells (Avilés-Jiménez et al., 2012[1]). The ability of Hp to cause disease is thought to attribute to complex interplay between host genetic factors, environmental and bacterial factors. H. pylori adheres to the cells of gastric epithelial and secretes effector molecules that can change gastric epithelial cell function and viability (Ernst and Gold, 2000[6]; Chatterjee et al., 2012[2]). These changes enhance production of cytokines, which are involved in gastric inflammation and epithelial cell damage (Avilés-Jiménez et al., 2012[1]). Moreover, Hp infection elicits a noticeable mucosal accumulation of T lymphocytes, which is responsible to gastric pathology by synthesizing T helper (Th) 1-type cytokines, such IFN-γ and TNF-α (D'Elios et al., 1997[4]; Ernst and Gold, 2000[6]; Hitzler et al., 2012[13]). Recently, a novel pathway of inflammation characterized by excessive production of IL-17 has been reported to be involved in the pathogenesis of immune-mediated diseases, such as rheumatoid arthritis, experimental autoimmune encephalomyelitis, and psoriasis (Kolls and Linden, 2004[15]; Steinman, 2007[33]). IL-17 consists of a family of related cytokines (IL-17A-F), IL-17- producing cells termed Th17 cells. These cells also produce TNF-α, IL-6, IL-22, and granulocyte macrophage-colony stimulating factor (Kolls and Linden, 2004[15]; Park et al., 2005[26]; Steinman, 2007[33]; Weaver et al., 2007[37]). IL-17 stimulates the synthesis of IL-1β, IL-6, TNF-α, PGE2, ICAM-1 and cyclo-oxygenase-2 (Fossiez et al., 1998[8]; Griffin et al., 2012[12]; Serelli-Lee et al., 2012[31]). Thus, IL-17 seems to provide a link between T cell activation and inflammatory responses. In spite of marked Th (Placeholder1) cell response, Hp-colonized gastric mucosa also contains high levels of IL-17 (Luzza et al., 2000[20]; Mizuno et al., 2005[22]). The synthesis of IL-8 by gastric mononuclear and epithelial cells is positively regulated by IL-17, thus emphasizing the potential role of IL-17 in the Hp-driven inflammation (Luzza et al., 2000[20]). However, the factors involved in the control of IL-17 production in Hp-associated gastritis are yet unclear.
IL-23 is a heterodimeric protein that is composed of a specific IL-23p19 subunit and IL-12p40 subunit. This cytokine is primarily produced by activated dendritic cells, monocytes and macrophages (Oppmann et al., 2000[24]). IL-23/p19-null mice have reduced numbers of Th17 cells, thus confirming the role of IL-23 in enhancing IL-17 production (Ghilardi et al., 2004[10]). The clinical outcome of H. pylori infection is suggested to be linked to certain strains such as the cytotoxin-associated gene (cagA) and vacuolating cytotoxin (vacA). The cagA gene which is a marker for the presence of a Pathogenicity Island (CagPAI), has been shown to be involved in induction of proinflammatory chemokine released.
The vacA gene has a mosaic structure and comprised signal (s) and middle (m) regions (Stathis et al., 2009[32]). The s region consists of any one of four signal sequences (type s1a, s1b, s1c or s2), while the m region consists of two middle region alleles (type m1 or m2) (Stathis et al., 2009[32]). The s1 subtype and m1 subtype have been linked to more severe clinical manifestation (Dong et al., 2008[5]; Ferrand et al., 2008[7]; Kuo et al., 2008[18]; Stathis et al., 2009[32]). CagA is present in more than 50 % of the H. pylori strains and encodes the cagA protein. It is a marker of the presence of the Pathogenicity Island, (Swisher and Barbati, 2007[35]; De Luca et al., 2008[3]) is related to virulence of the H. pylori strain and is associated with peptic ulcer and gastric malignancy in some populations. This study was aimed to evaluate the association of the virulence factors of Hp and gastric mucosal interleukin-17/23 mRNA expression in dyspeptic patients.
Materials and Methods
A total of 58 H. pylori-infected gastritis patients 23 men (43.43 ± 3.53) and 35 women (40.57 ± 2.44) were participated in this study. Hp infection was determined by the rapid urease test, PCR 16srRNA, urea and histological examination of biopsies taken from the corpus. Patients were classified as Hp-infected only if the three tests were positive, respectively. Four biopsies were collected from 58 Hp-infected, and used for rapid urease test, histological examination, assessment of bacterial virulence factors, detection of H. pylori and cytokine RNA analysis.
Histological examination
Sections of biopsy specimens were embedded 10 % buffered formalin and stained with hematoxylin and eosin to examine gastritis and with giemsa to detect Hp. The histological severity of gastritis was blindly graded from normal to severe based on the degree of mononuclear cell (MNC) and polymorphonuclear leukocyte (PMN) infiltration, and atrophy according to the Updated Sydney system (Manxhuka-Kerliu et al., 2009[21]) on a four-point scale: 0, no; 1, mild; 2, moderate; and 3, severe changes.
PCR amplification
DNA for polymerase chain reaction (PCR) was extracted using the Bioflux tissue (Bioflux, Japan). Oligonucleotide primers for PCR amplification of specific segments are shown in Table1(Tab. 1). For vacA and cagA evaluation, the PCR program comprised 35 cycles of denaturation (at 94 °C for 30 s), annealing (at 56 °C for 30 s, extension at 72 °C for 30 s), and one final extension (at 72 °C for 5 min).
Quantitative analyses for IL-17 and IL-23 mRNA in the gastric mucosa using real-time RT-PCR
Total RNA was isolated from whole gastric biopsy specimens using total RNA extraction biozol (Bioflux, Japan). An aliquot containing 0.2 μg of total RNA was used for the reverse transcription reaction, which was conducted using the Superscript first-strand cDNA synthesis system (Fermentas, Finland) according to the manufacturer's instructions. The sequences of oligonucleotide primer and probe are shown in Table 2(Tab. 2). The quantification of IL-17 and IL-23 mRNA levels was performed using a Rotor-Gene 3000 (Corbett). Q-PCR reactions were performed in a total volume of 25 µl containing 3 µl of synthesized cDNA solution, 12.5 µl of 2x Rotor-Gene Probe PCR Master Mix (Qiagen, Germany), 500 nM of each primer and 250 nM of the TaqMan probe. Amplification program included a pre warming step (10 min at 94 ºC), denaturation step (94 °C for 15 s) and an annealing/extension step (60 °C for 60 s). β-actin was quantified as a reference gene to normalize the mRNA expression levels of other genes. The relative quantification of gene expression in each sample was analyzed by the 2ΔΔCt method and expressed as the ratio of related gene to β-actin mRNA.
Statistical analysis
All experiments were performed at least three times. Parametric data are presented as mean and SE and nonparametric data are presented as medians (25-75 % quartiles). Statistical analysis was performed by Mann-Whitney Rank Sum test or nonpaired t test depending on the data set. A p value of <0.05 was accepted as statistically significant.
Results
Effect of virulence factors in H. pylori-infected on the Mucosal IL-17 and IL-23 mRNA levels in gastric mucosa
In our study, no significant differences were observed between the expression of interleukin 17 and 23 in Helicobacter pylori positive and negative patients (Figure 1(Fig. 1)). Our results showed that in H. pylori infected patients; mucosal IL-17 and IL-23 mRNA levels were dependent on the vacA (m1 and m2) status. Mucosal IL-17 and IL-23 mRNA expression in gastritis patients with vacA (m1)-positive were significantly higher than those observed in gastritis patients with vacA (m2) - positive (Figure 2(Fig. 2)). Mucosal IL-17 and IL-23 mRNA expression in gastritis patients with vacA (s1m1)-positive was also significantly higher than those observed in gastritis patients with vacA (s1m2)-positive (Figure 3(Fig. 3)). Also no significant differences were observed between the expression of interleukin 17 and 23 in gastritis patients with cagA-positive and cagA-negative (Figure 4(Fig. 4)).
Correlation between virulence factors and gastric mucosal inflammation and types of disease
Severe polymorphonuclear infiltration and chronic gastritis were significantly higher in gastritis patients with vacA (m1)-positive compare to those observed in gastritis patients with vacA (m2)-positive. Severe polymorphonuclear infiltration and chronic active gastritis were also significantly higher in gastritis patients with cagA-positive compared to those observed in gastritis patients with cagA-negative patients (Table 3(Tab. 3)).
Correlation between Mucosal IL-17 and IL-23 mRNA levels and gastric mucosal inflammation and types of disease
In those infected with H. pylori, IL-17 and IL-23 mRNA expression in mucosa did not correlate with the degree of mononuclear (P=0.192 and P=0.471) or polymorphonuclear (P=0.279 and P=0.628) cell infiltration, chronic gastritis (P=0.192 and P=0.471), or chronic active gastritis (P=0.279 and P=0.628) assessed by the Sydney system.
Discussion
In this study we aimed to evaluate the association of the virulence factors of Hp and gastric mucosal interleukin-17/23 mRNA expression in patients with gastritis. Our results showed that there was no significant difference between the expression of interleukin 17 and 23 in Helicobacter pylori positive and negative patients. That might be related to the presence of gastritis in two groups. The results of the present study showed that mucosal IL-17 and IL-23 mRNA expression were dependent on the vacA (m1) status and their expression in patients infected with vacA m1-positive strains were significantly higher than those observed in patients with vacA m2-positive strains. Until now a similar study has not been done and this is the first report about correlation of vacA with IL-17 and IL-23 expression.
In this study, we determined that the expression of interleukin 17 and 23 in all patients was high. It has been reported that the IL-23-IL-17 axis plays an important role in the development of chronic inflammation and in host defenses against bacterial infection (Gisbert and Calvet, 2011[11]; Kuo et al., 2011[17]; Nakamura et al., 2012[23]). Also vacuolating activity is higher in s1/m1 genotypes than in s1/m2 genotypes, and is absent in s2/m2 genotypes (Ota et al., 2009[25]; Suzuki et al., 2009[34]; Park et al., 2010[27]). Therefore, it can be concluded that vacA s1/m1 strains cause more immune response and are more frequently associated with gastritis.
Considering that expression of interleukins 17 and 23 in patients with vacA m1 exists in chronic gastritis, therefore, the overexpression of these interleukins along with vacA m1 may similarly lead to chronic gastritis. In our study, the relationships between cagA-positive and severe polymorphonuclear infiltration and chronic active gastritis were significantly higher than those observed in patients with cagA-negative. While in Western countries it has been demonstrated that more severe gastric inflammation develops after infection with cagA-positive strains and in Asian countries there is no such difference (Umit et al., 2009[36]; Rathbone and Rathbone, 2011[29]).
It has been reported that in Western countries, cagA-positive status has been associated with the severe outcomes of the infection (Kondo et al., 2009[16]; Lehours et al., 2009[19]; Zullo et al., 2009[38]). It is possible that the relationship between H. pylori genotypes and infection with gastric inflammatory response varies in different nations. These variations in the clinical consequences are due to factors such as duration of the infection, inflammatory response of the patient, virulence of H. pylori strains, etc. Although our results do not support the correlation of IL-17 and IL-23 with the inflammatory phenotype of H. pylori infected patients, but emerging experimental evidence suggests that IL-23/IL-17 pathway is an important dynamic force for existing gastric inflammation in H pylori-infected patients. However, further studies are needed to establish the exact contribution of each of these cytokines in the H pylori-associated gastric pathology.
References
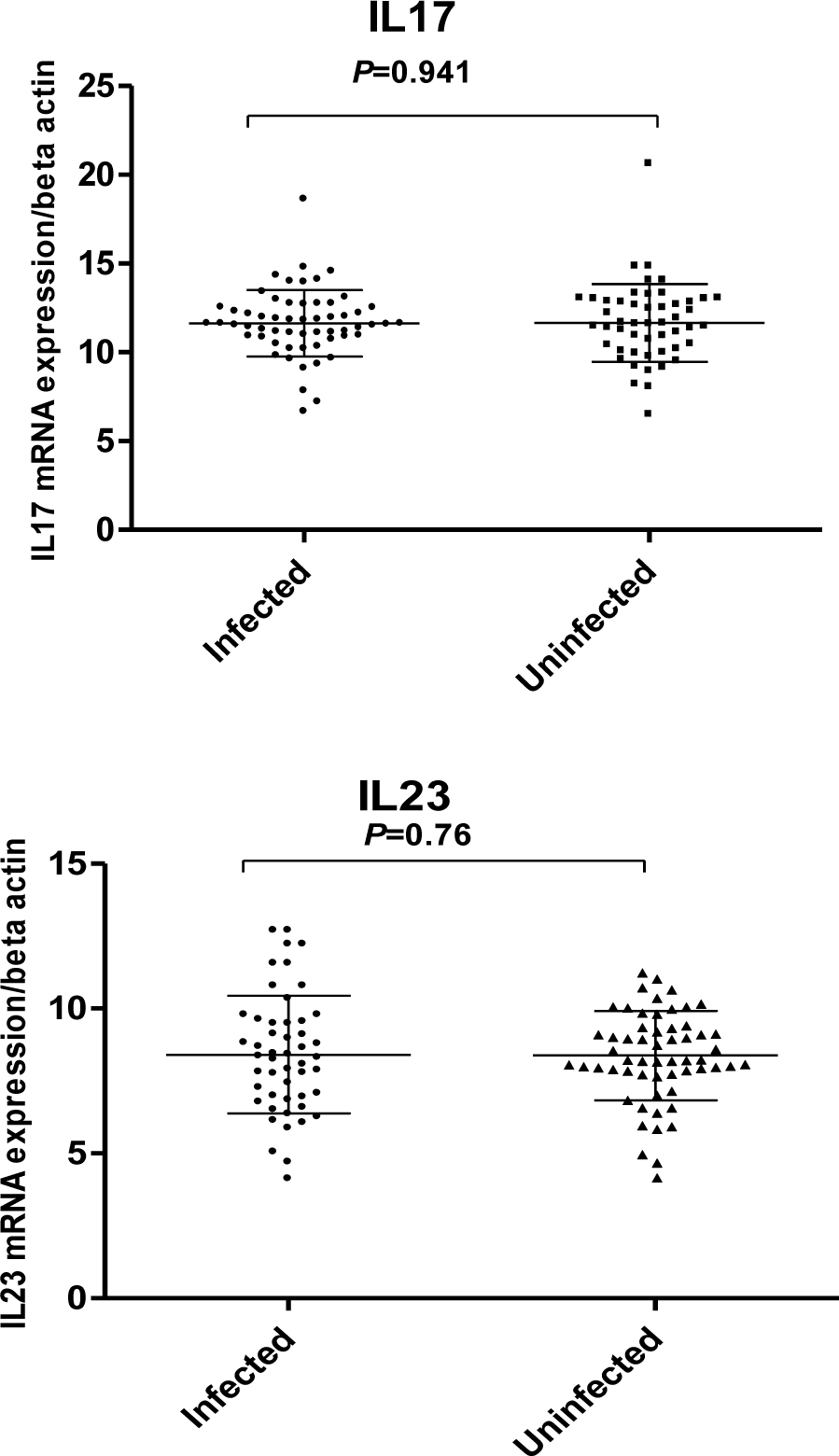
Figure 1: Mucosal IL-17 and IL-23 mRNA expression in gastritis patients. RNA was extracted from gastric biopsies of 58 Hp-infected patients, 50 Hp non-infected patients with gastritis and analyzed for IL-17 and IL-23 by real time PCR. Levels are normalized to β-actin. Values are presented as mean ± SD of all experiments.
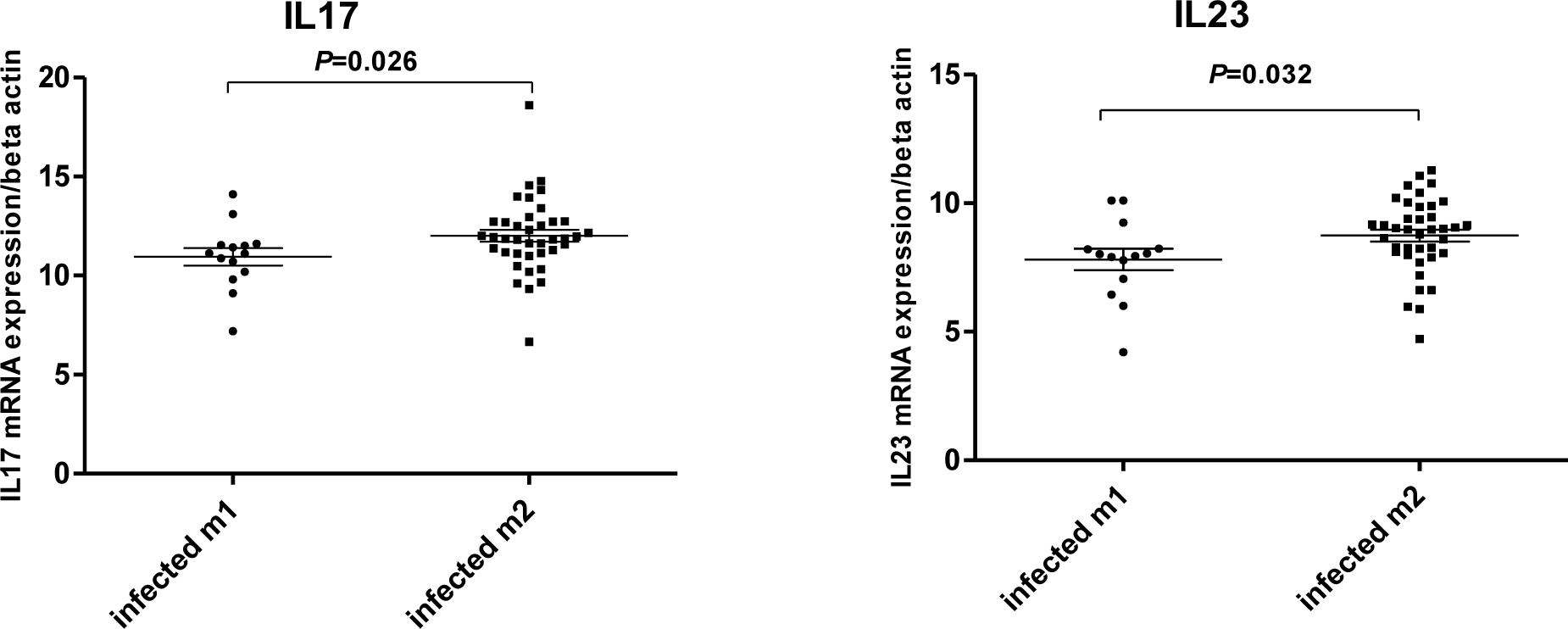
Figure 2: Mucosal IL-17 and IL-23 mRNA expression in gastritis patients with vacA-positive. RNA was extracted from gastric biopsies of 14 Hp-infected patients with vacA (m1)-positive, 30 Hp-infected patients with vacA (m2)-positive with gastritis and analyzed for IL-17 and IL-23 by real time PCR. Levels are normalized to β-actin. Values are presented as mean ± SD of all experiments.
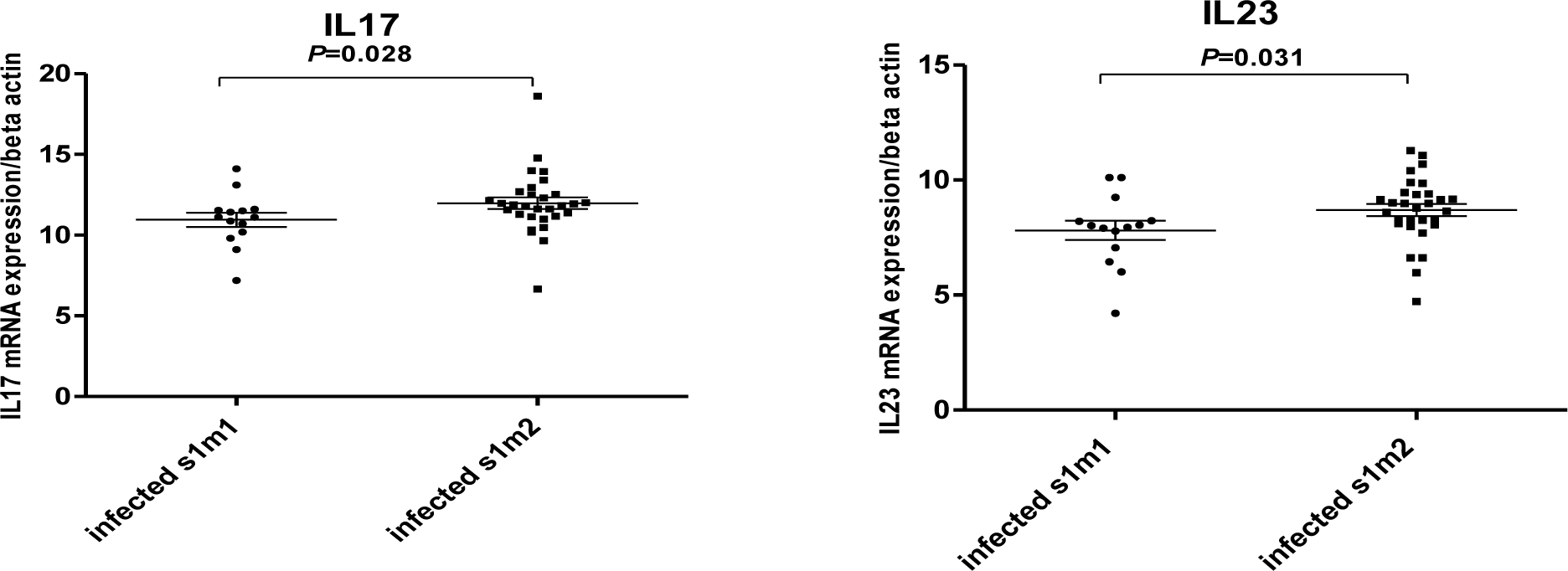
Figure 3: Mucosal IL-17 and IL-23 mRNA expression in gastritis patients with vacA-positive. RNA was extracted from gastric biopsies of 14 Hp-infected patients with vacA (s1m1)-positive, 30 Hp-infected patients with vacA (s1m2)-positive with gastritis and analyzed for IL-17 and IL-23 by real time PCR. Levels are normalized to β-actin. Values are presented as mean ± SD of all experiments.
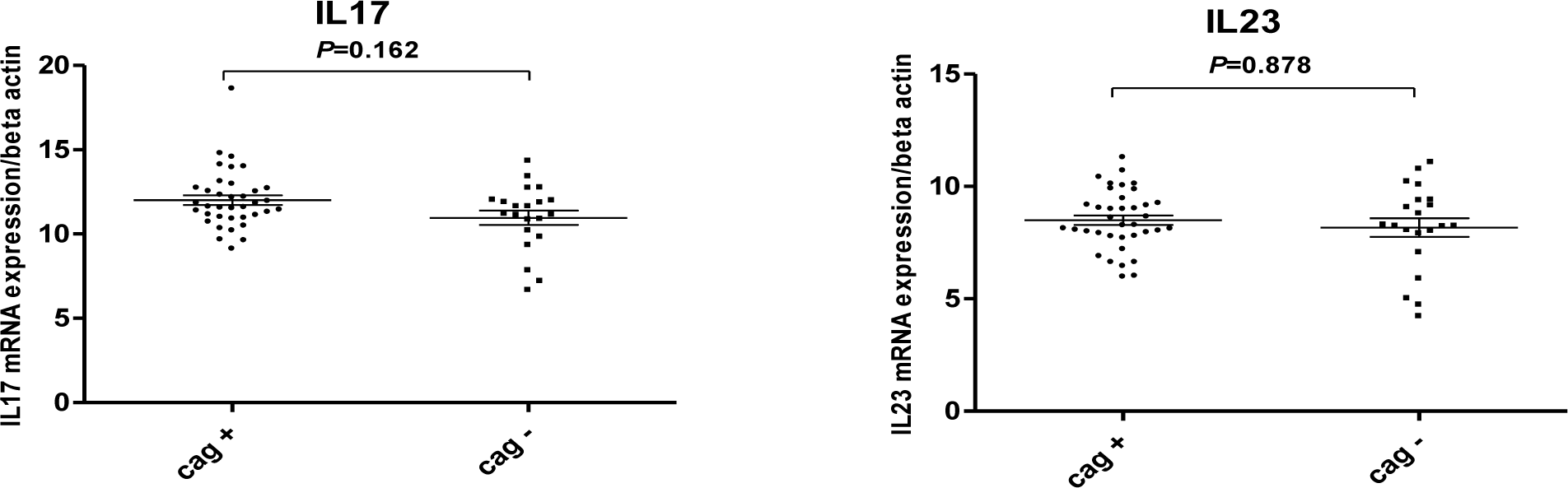
Figure 4: Mucosal IL-17 and IL-23 mRNA expression in Hp-infected patients with cagA-positive and cagA-negative. RNA was extracted from gastric biopsies of 37 Hp-infected patients with cagA-positive, 21 Hp-infected patients with cagA-negative and analyzed for IL-17 and IL-23 by real time PCR. Levels are normalized to β-actin. Values are presented as mean ± SD of all experiments.


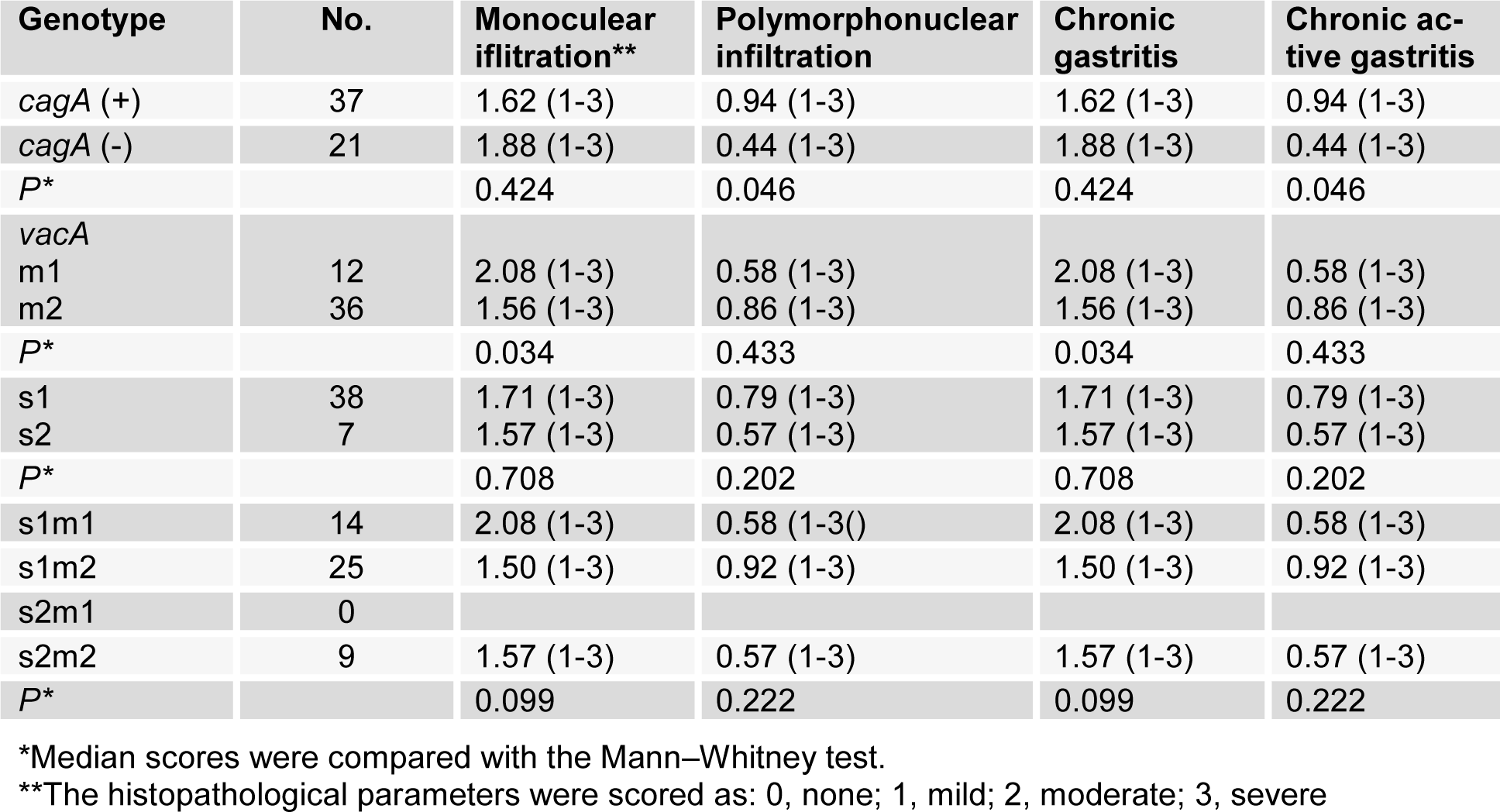
Table 3: Relationship between histological parameters determined in gastric biopsy specimens and virulence factors
[*] Corresponding Author:
Hedayatollah Shirzad, Cellular and Molecular Research Center, Shahrekord University of Medical Sciences, Shahrekord, Iran, eMail: shirzad1951@yahoo.com or shirzadeh@SKUMS.ac.ir