Research article
Seasonal variability of Chelidonium majus L. secondary metabolites content and antioxidant activity
Z. Dragana Jakovljevic1[*], S. Milan Stankovic1, D. Marina Topuzovic1
1Department of Biology and Ecology, Faculty of Science, University of Kragujevac,str. Radoja Domanovica No. 12, 34000 Kragujevac, Republic of SerbiaEXCLI J 2013;12:Doc260
Abstract
The aim of this study is to investigate the total phenolic content, concentration of flavonoids and antioxidant activity in extracts of the plant Chelidonium majus L. during different phenological stages (stage of rosette, the initial flowering stage, the stage of fully formed flowers and stage of fruits formation). Five different extracts of the whole plant, for each phase, were obtained by extraction with water, methanol, acetone, ethyl acetate and petroleum ether. The concentration of total phenolic content was determined using the Folin-Ciocalteu´s reagent and obtained values were the highest in the rosette stage (60.96 mg GA/g). The concentration of flavonoids was the highest in the initial stage of flowering (291.58 mg RU/g). The antioxidant activity was determined in vitro using DPPH reagent. The highest antioxidant activity was expressed in the rosette stage (50.72 mg/ml). Based on the obtained results it can be concluded that the concentrations of secondary metabolites in Ch. majus depend on the phenological stage of the plant.
Keywords: Ch. majus, seasonal variation, antioxidant activity, phenols, flavonoids
Introduction
Chelidonium majus L. (Papaveraceae) is a perennial herbaceous plant, 50-100 cm of height, with an upright and spreading stem, large leaves and yellow flowers collected on the tops of the stems in rare umbel inflorescence. The plant is widely present in Europe and Asia, growing at neglected rocky positions near villages. For centuries, it has been used in folk medicine as a keratolytic and keratoplastic. All parts of the plant contain orange latex rich in alkaloids, among which the most present are chelidonine, chelerythrine, sanguinarine, berberin and others (Colombo and Bosisio, 1996[4]). Numerous studies show the high content of bioactive components with antiparasitic (Yao et al., 2011[23]), anti-inflammatory (Lee et al., 2007[10]), antifungal (Meng et al., 2009[11]), antimicrobial (Kokoska et al., 2002[8]), and cytotoxic effects (Nadova et al., 2008[13]; Kaminsky et al., 2006[7]).
Due to their notable pharmacological effects, Ch. majus is widely used in traditional and modern medicine for the treatment of liver diseases, gastrointestinal tract, and there are also some data on the use of this herb for the prevention and treatment of cancer and tumors (Venkatesh et al., 2011[21]; de Melo et al., 2011[5]).
Fundamental metabolic processes of plants are considered to be the primary metabolic processes that occur by the same mechanism in the cells of all plants. These chemical processes must be produced by each plant on a daily basis in order to survive and to reproduce. However, the plants produce a large number of compounds, secondary metabolites, which enable the biochemical communication inside the ecosystem. Biochemical aspect of the synthesis of secondary metabolites depend on the plant genetic, taxonomy, the stage of development, the season, the presence of parasites and others. The variations could also be the result of abiotic factors such as relief, altitude, geological substrate characteristics, etc.
Secondary metabolites and their derivatives show significant biological and pharmacological properties, such as hepatoprotective, diuretic, spasmolytic. They also show antioxidant, antiallergic and anticancer effects (Williams et al., 2004[22]; Mulubagal and Tsay, 2004[12]; Borneo et al., 2008[3]).
Maria Laura Colombo and Enrico Bosisio (Colombo and Bosisio, 1996[4]) investigated the pharmacological activity of greater celendine and apart from identifying antiviral, antitumor and antimicrobial activities, they also identified the presence of flavonoids and phenolic acids. The application of FRAP method and DPPH reagent in alcoholic extracts of this plant showed significant antioxidant activity (Then et al., 2003[20]; Nadova et al., 2008[13]).
According to the literature data, Ch. majus has many important biological properties, but there is little data about whether the biological properties of these plants are different during its growing season. The main objective of this study was to determine how the phenolic content, flavonoid concentration and antioxidant activity in different types of plant extracts Ch. majus vary depending on the phenological phases of plants, as well as to determine at what stage is the greatest concentration of these secondary metabolites.
Materials and methods
Plant material
Aboveground plant parts Ch. majus were collected at sites in Šumarice, Kragujevac, central Serbia, during the period from April to August, 2010. In April the plant was not sufficiently developed, rosette leaves were formed but the flowers were not formed yet. At the end of May the collected samples were in the second phenological phase. At this stage the plant had the upright stem, fully formed leaves, and was in early flowering stage (buds were formed). In early July, the samples were in the third phase with a clearly formed inflorescence. The last phenological phase of collected samples was in August. The plant was passe and the fruit was in the process of forming.
Chemicals
Acetone, methanol, petroleum ether, ethyl acetate and sodium hydrogen carbonate were purchased from "Zorka pharma" Šabac, Serbia. Standards of phenolic acids (gallic acid) and flavonoids (rutine hydrate), chlorogenic acid and 2,2-diphenyl-1-picrylhydrazyl (DPPH) were obtained from Sigma Chemicals Co. manufacturers., St. Louis, MO, USA. Folin-Ciocalteu's phenol reagent, 3-tetra-butyl-4-hydroxyanisole (BNA) and aluminum chloride (AlCl3) were from Fluka Chemie AG, Buchs, Switzerland.
Preparation of plant extracts
Sampled plant material was dried at 20 °C in a dark place. After drying, the material was fragmented. 2 g for each phenological phase was extracted with 50 ml of solvent (water, methanol, ethyl acetate, acetone and petroleum ether) using a Soxhlet apparatus. After 24 h, infusions were filtered through Whatman No.1 filter paper and residue was re-extracted with equal volume of solvents. After 48 h, the process was repeated. Combined supernatants were evaporated to dryness under vacuum at 40 °C using Rotatory evaporator. The obtained extracts were kept in sterile sample tubes and stored in a refrigerator at 4 °C.
Determination of total phenolic contents in plant extracts
The concentration of phenolics in plant extracts was determined using spectrophotometric method (Singleton et al., 1999[16]). Methanolic solution of the extract in the concentration of 1 mg/ml was used in the analysis. The reaction mixture was prepared by mixing 0.5 ml of methanolic solution of extract, 2.5 ml 7.5 % NaHCO3. Blank was concomitantly prepared, containing 0.5 ml methanol, 2.5 ml 10 % Folin-Ciocalteu´s reagent dissolved in water and 7.5 % NaHCO3. The samples were incubated at 45 °C for 15 min. The absorbance was determined at λmax = 765 nm. The samples were prepared in triplicate and the mean value of absorbance was obtained. Blank was concomitantly prepared with methanol instead of extract solution. The same procedure was repeated for the gallic acid and the calibration line was construed. The total phenolic content was expressed in terms of gallic acid equivalent (mg of GA/g of extract).
Determination of flavonoid concentrations in the plant extracts
The concentration of flavonoids was determined using spectrophotometric method (Quettier et al., 2000[15]). The sample contained 1 ml of methanolic solution of the extract in the concentration of 1 mg/ml and 1 ml of 2 % AlCl3 solution dissolved in methanol. The samples were incubated for an hour at room temperature. The absorbance was determined at λmax = 415 nm. The samples were prepared in triplicate and the mean value of absorbance was obtained. The same procedure was repeated for the rutin and the calibration line was construed. The concentration of flavonoids in extracts was expressed in terms of rutin equivalent (mg of RU/g of extract)
Evaluation of antioxidant activity
The ability of the plant extract to scavenge DPPH free radicals was assessed using the method described by Tekao et al. (1994[19]), adopted with suitable modifications from Kumarasamy et al. (2007[9]). The stock solution of the plant extract was prepared in methanol to achieve the concentration of 1 mg/ml. Dilutions were made to obtain concentrations of 500, 250, 125, 62.5, 31.25, 15.62, 7.81, 3.90, 1.99, 0.97 µg/ml. Diluted solutions (1 ml each) were mixed with 1 ml of DPPH methanolic solution (80 µg/ml). After 30 min in darkness at room temperature (23 °C) the absorbance was recorded at 517 nm. The control samples contained all the reagents except the extract. The percentage inhibition was calculated using the equation: % inhibition = 100 x (A control - A sample)/A control), whilst IC50 values were estimated from the % inhibition versus concentration sigmoidal curve, using a non-linear regression analysis. The data were presented as mean values ± standard deviation (n = 3).
Results
In order to determine the seasonal variability of the concentration of phenolic compounds, flavonoid concentration and antioxidant activity, the analysis of plant material, sampled during four different phenological stages of Ch. majus, within the period from April to August 2010, was performed. Five extracts were obtained by extraction with water, acetone, methanol, ethyl acetate and petroleum ether, for each phenological stage.
Total phenolic content
The results of total phenolic content of the examined plant extracts, determined using Folin-Ciocalteu's method, are presented in Table 1(Tab. 1). The values of total phenolic content in the extracts are expressed as gallic acid equivalents.
The obtained values for the first phenological stage ranged from 16.45 to 60.96 mg GA/g. In the second phase, the obtained values ranged from 12.72 to 38.99 mg GA/g. For the third phenological stage, the obtained values were of 5.74 to 38.7 mg GA/g. The highest concentration of total phenolic compound for all three phases was gained in the methanol extract. The fourth stage is characterized by the concentration of total phenolic compounds in the range from 10.39 to 48.48 mg GA/g. At this stage, the highest value was obtained in the acetone extract.
By analyzing the results of total phenolic content in all extracts for all plant stages, it was noticed that the highest concentration of total phenolic compounds were obtained from plants collected during the month of April when the plant is in the rosette stage (60.96 mg GA/g). In the period from May to July, the concentration of phenolic compounds was decreasing (the lowest value in the flowering stage, and the aqueous extract was 5.74 mg GA/g), to increase again in August, when it initiated the formation of fruit. Methanol is the most effective solvent for extraction of phenolic compound from Ch. majus. The highest concentration of phenolic compounds in the extracts were obtained using the solvents of moderate polarity.
Flavonoid concentration
The concentration of flavonoids in various extracts of Ch. majus, collected during different stages, was determined using spectrophotometric method with AlCl3. The content of flavonoids was expressed in terms of rutin equivalents. The summary of quantities of flavonoids identified in the tested extracts is shown in Table 2(Tab. 2).
The first stage extracts showed concentration of flavonoids in the range of 7.12 to 233.89 mg RU/g, the second phase extracts of 10.44 to 291.58 mg RU/g, and the third from 7.44 to 135.37 mg RU/g. For all three phenological phases the highest concentrations of flavonoids were obtained in acetone extracts. In the fourth stage, the flavonoid concentrations ranged from 8.53 to 242.30 mg RU/g, with the highest concentration achieved in the ethyl acetate extract.
The highest concentration of flavonoids (291.58 mg RU/g) was recorded during the month of May, in the second phase, just before flowering. After the completion of this phase, during flowering, the decline in the concentration of flavonoids was observed (the lowest concentration of the aqueous extract of the rosette stage and flowering stage - 7.12 and 7.44 mg RU/g), and then at the beginning of the formation of flowers, flavonoid concentrations again increased. Acetone and ethyl acetate are the most suitable solvents for the extraction of this type of extract.
Antioxidant activity
The antioxidant activity of different plant extracts of Ch. majus, collected during different stages, was determined using methanol solution of 2,2-diphenyl-1-picrylhydrazyl (DPPH) reagent. DPPH method was also used to quantify antioxidants in complex biological systems in recent years and it's based on the reduction of methanolic solution of colored free radical DPPH by free radical scavenger. The scavenging activity was measured as the decrease in absorbance of the samples versus DPPH standard solution. The results are shown in Table 3(Tab. 3).
In the rosette stage antioxidant value ranged from 50.72 to 858.33 mg/ml. The second phase is characterized by a range of 68.05 to 255.21 mg/ml. The highest values obtained for the third and fourth phases are 196.56 mg/ml and 192.99 mg/ml. For each of the stages the highest values were obtained in methanolic extracts.
Based on these results, the highest DPPH radical neutralizing abilities were manifested in methanol extract of the plant during the rosette stage, which at a concentration of 50.72 mg/ml neutralizes 50 % of free radicals. Approximate activity was manifested in methanol extract of the stage before flowering (IC50 value was 68.05 mg/ml). The minimum capability of DPPH radical inhibition was recorded for the petroleum ether extract of the flowering stage and for the water extract of the flowering stage, and the IC50 values for these extracts were not determined.
Discussion
The ability of the synthesis of secondary metabolites depends on the direction of plant ontogeny. Each plant species enters its specifities into metabolic processes which result in the synthesis of various metabolites. Variations in the concentration of secondary metabolites are the result of both biotic and abiotic factors. Quantitative and qualitative analysis of the dynamics and distribution of secondary metabolites, the impact of environmental factors on their structure as well as biological activity, provide answers to many current issues in several disciplines (Stanković et al., 2010[18]; Stanković et al., 2011[17]).
Phenols are a large class of secondary metabolites. Phenolics (including flavonoids) are among most active antioxidants, as well as most important stabilization factors of the oxidative processes. Flavonoids include many pharmacological and biological properties. In phytopharmacy flavonoids are the active ingredients, of plant origin, with strong biological effects, while in dietetics and food industries they present important phytonutrients, preservatives, spices and aromatics substances (Stanković et al., 2010[18], 2011[17]). They also present the objects of researches both from the aspects of adaptation of plants to various biotic and abiotic factors, and of the interaction of plants in natural ecosystems.
In order to determine the seasonal dynamics of the concentration of secondary metabolites in plant species Ch. majus, the material was sampled during four phenological phases, and the results indicate that there is a significant fluctuation of secondary metabolites during the life cycle of these species. The highest concentration of total phenolic compounds was observed in the spring, when the plant was in the rosette stage with well-formed, thick leaves, but when the whole plant is not yet sufficiently developed. When the plant enters the flowering stage, there is a lack of that concentration, which again increases when the plant begins with the formation of fruit. The concentration of flavonoids is the greatest just before flowering and before fruiting. During flowering the concentrations of these secondary metabolites is the lowest. The antioxidant activity and total phenol concentrations is the highest in the spring months during the rosette stage.
Papageorgiou et al. (2008[14]) studied the seasonal variation of total phenolics, antioxidant activity, and flavonoids in plant extracts Rosmarinus officinalis and Salvia fruticosa during three phenological stages (before flowering, during flowering and fruiting in). Extracts of rosemary and sage showed the highest concentration of total phenols in the period before and during flowering when the fruits are developed. In the flowering stage, according to these authors, the concentration of total phenols is the lowest. The antioxidant activity was correlated with the concentration of total phenolics. Flavonoids showed the greatest activity during flowering. According to these authors, the combination of environmental factors such as temperature, humidity, rainfall, etc. can speed up or slow down the accumulation of phenolics present in the plants.
Yasar et al. (2010[24]) studied the seasonal variation of total phenolic compounds, antioxidants and minerals in the fresh shoots of green tea (Camellia sinensis var. sinensis) in May, July and September. According to results of this study, antioxidant activity and total phenol concentrations increase from May to July, that is, the highest values were recorded in the warmest month. The differences in the concentration of total phenolic compounds, these authors explain according to the differences between day and night temperatures, but also to the length and strength of the sunlight. The differences in antioxidant activities within the season are due to various external factors during these three months.
Numerous modern studies have shown that in response to environmental stimuli plants produce adequate secondary metabolites. Biochemical behavior, as a respond to stimuli, allows plants quickly to respond and recover from the environmental stress, as well as to adapt to the environment. Synthesis and accumulation of phenolics is regulated by a complex interaction between internal factors (phylogeny and ontogeny) and environmental factors - both abiotic (light, nutrient availability) and biotic (other enemy) (Beckman, 2000[1]; Booij-James et al., 2000[2]).
During ontogenetic development, each individual plant species is exposed to environmental factors. The answer will depend on the ability of the plant itself. Harmful effects of the distortion of antioxidant-prooxidant balance of plants can largely prevent the synthesis of the corresponding metabolites. The period in which the plant blossoms and bears fruit is the most important in terms of plant species survival.
The obtained values for the concentration of total phenolic compounds indicate that the highest concentration is during April, when the plant is in the rosette stage, and then, in the stage fruit formation. The highest concentration of flavonoids is just before flowering and it also increases in the fruit formation stage. The highest DPPH radical neutralizing abilities manifested methanol extract of the plant in the rosette stage, and approximate activity manifests and methanol extract of the stage before flowering.
Neither strong nor direct relations were found between the antioxidant activity of the plant and the concentrations of flavonoid content and phenolic compounds. This could be due to the complexity of the substances which cause the antioxidant activity of this plant. The composition evolution in two opposite ways simultaneously for two compounds characterized by high individual activities could lead to the compensatory effect of the final activity (Gourine et al., 2010[6]).
In order to determine the components responsible for the high activity of Ch. majus, these authors suggest that another study must be conducted which would aim the isolation of components with antioxidants activity in rossete stage.
Conclusion
This research shows that there is a major difference in total phenolic content, flavonoids concentration and antioxidant activity during the growing season of Ch. majus, and that they depend on the phenological stage in which the plant is. Furthermore, their compositions vary in different extracts of this plant. It is noticed that the highest concentration is obtained using solvents of high polarity.
Therefore, information obtained here can be useful to get a higher concentration of these substances since this species has a long history of commercial use.
Acknowledgements
This work was supported by the Ministry of Science and Techological Development of the Republic of Serbia (III41010).
References
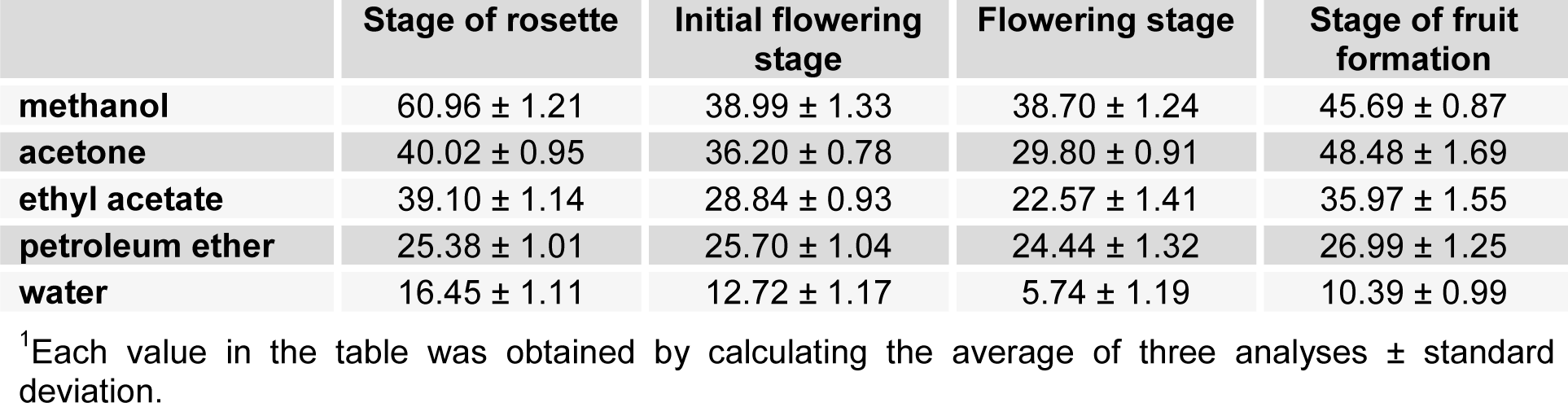
Table 1: Total phenolic contents1 of Ch. majus extracts expressed in terms of gallic acid equivalent (mg of GA/g of extract)
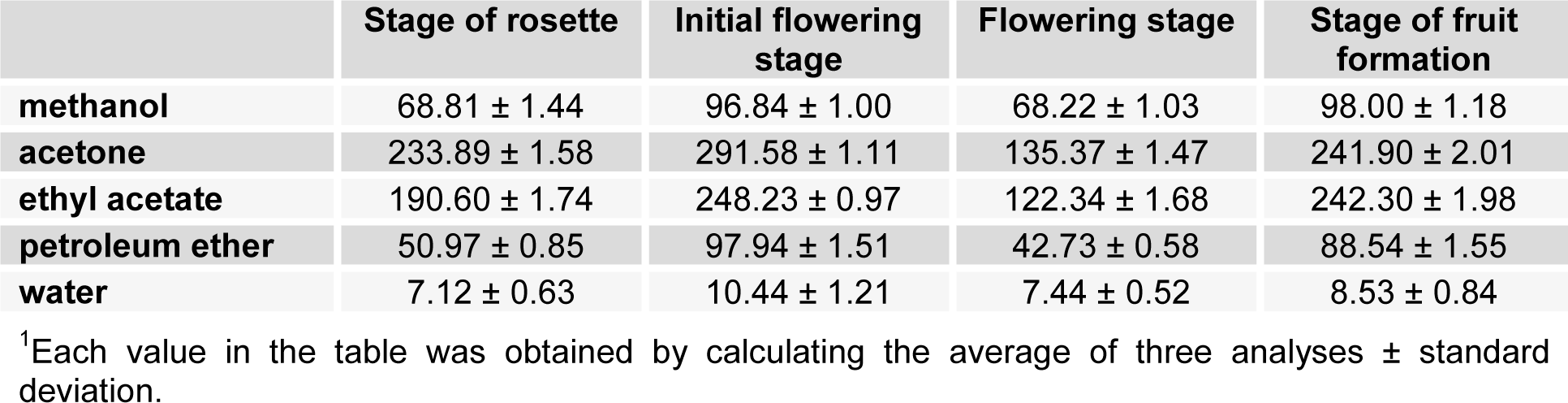
Table 2: Flavonoid concentrations1 of Ch. majus extracts expressed in terms of rutin equivalent (mg of RU/g of extract)
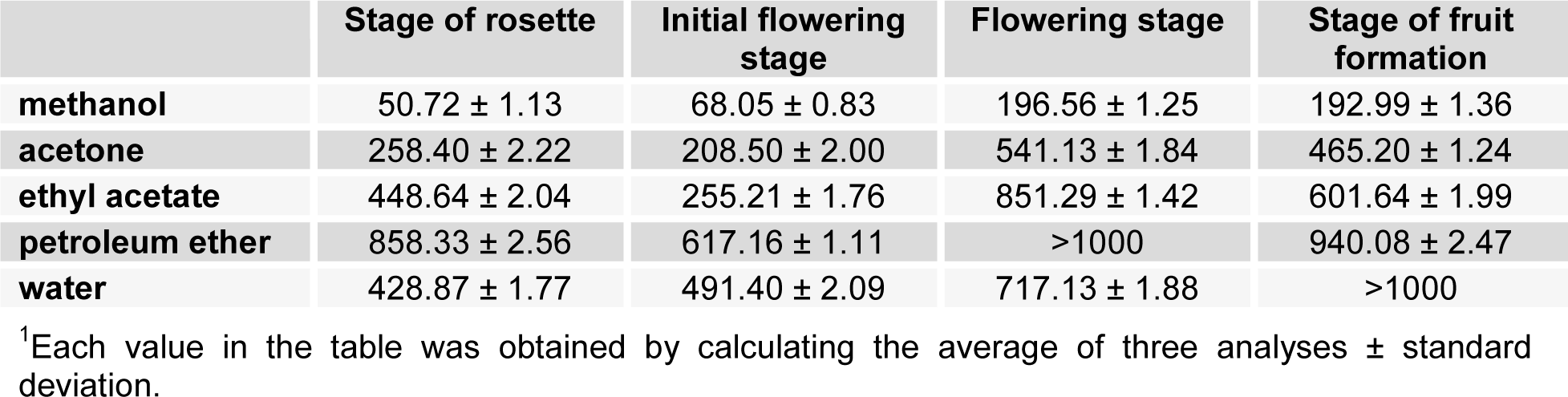
Table 3: Antioxidant (DPPH scavenging) activity1 of Ch. majus extracts presented as IC50 values (µg/ml)
[*] Corresponding Author:
Z. Dragana Jakovljevic, Department of Biology and Ecology, Faculty of Science, University of Kragujevac,str. Radoja Domanovica No. 12, 34000 Kragujevac, Republic of Serbia; Tel.: +381 34 336 223; Fax: +381 34 335 040, eMail: draganajakovljevic@ymail.com