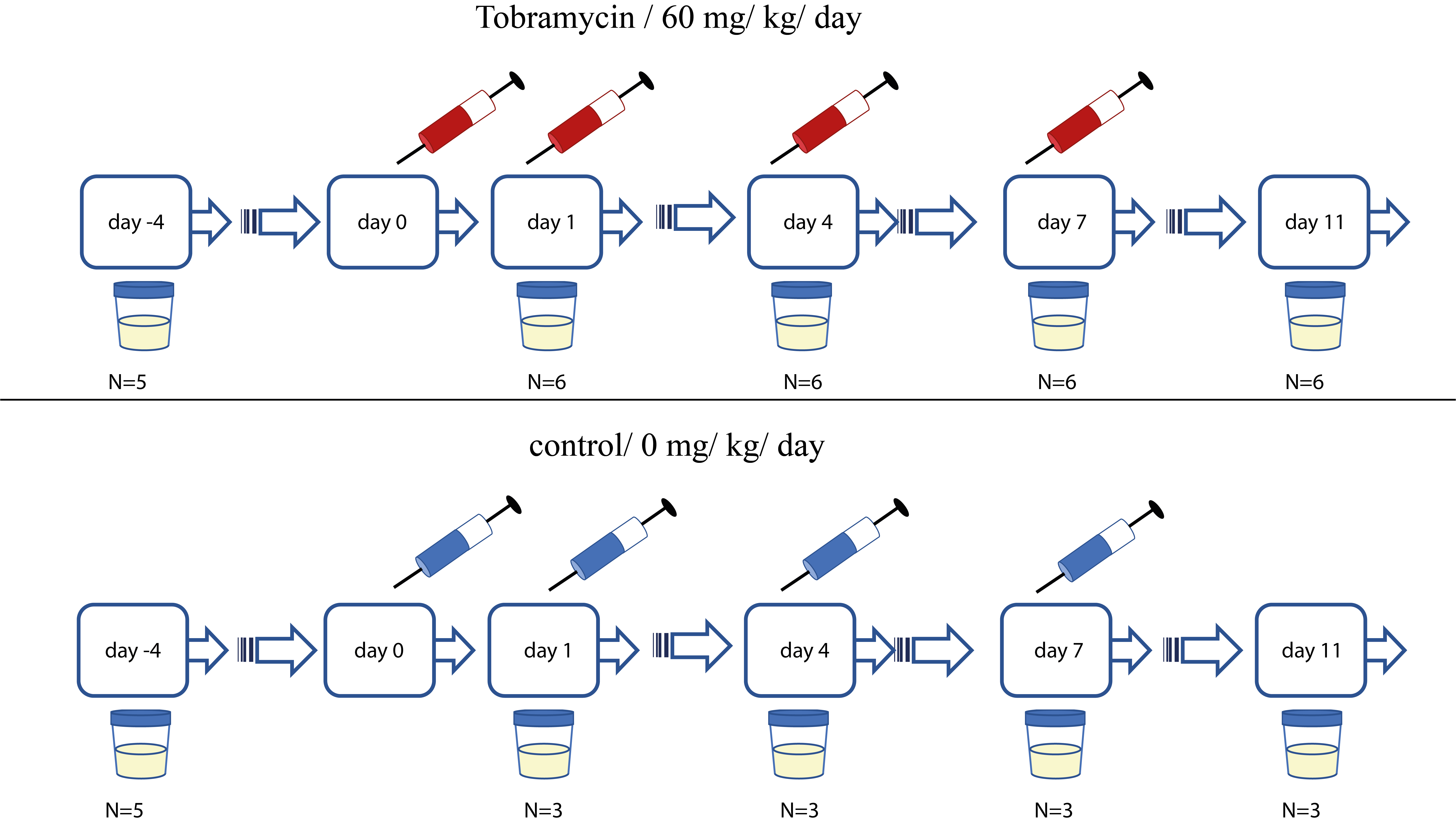
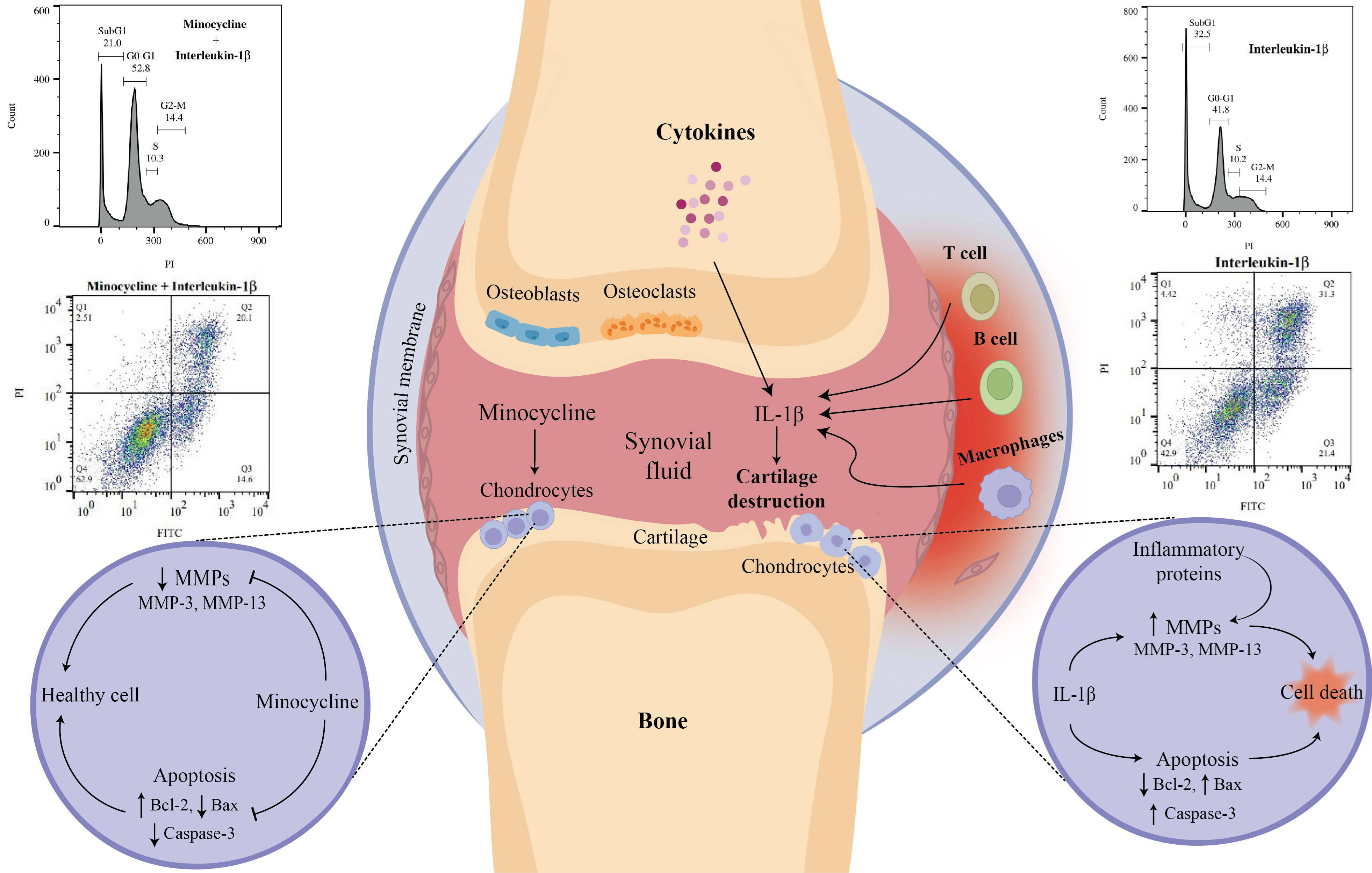
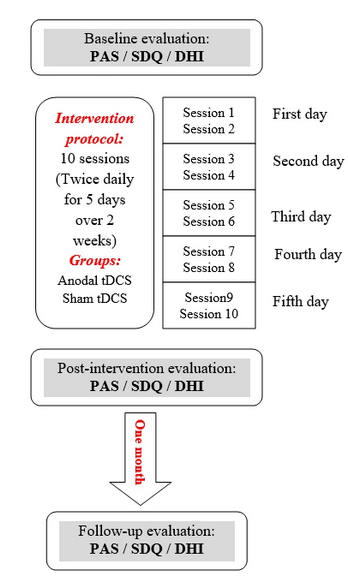
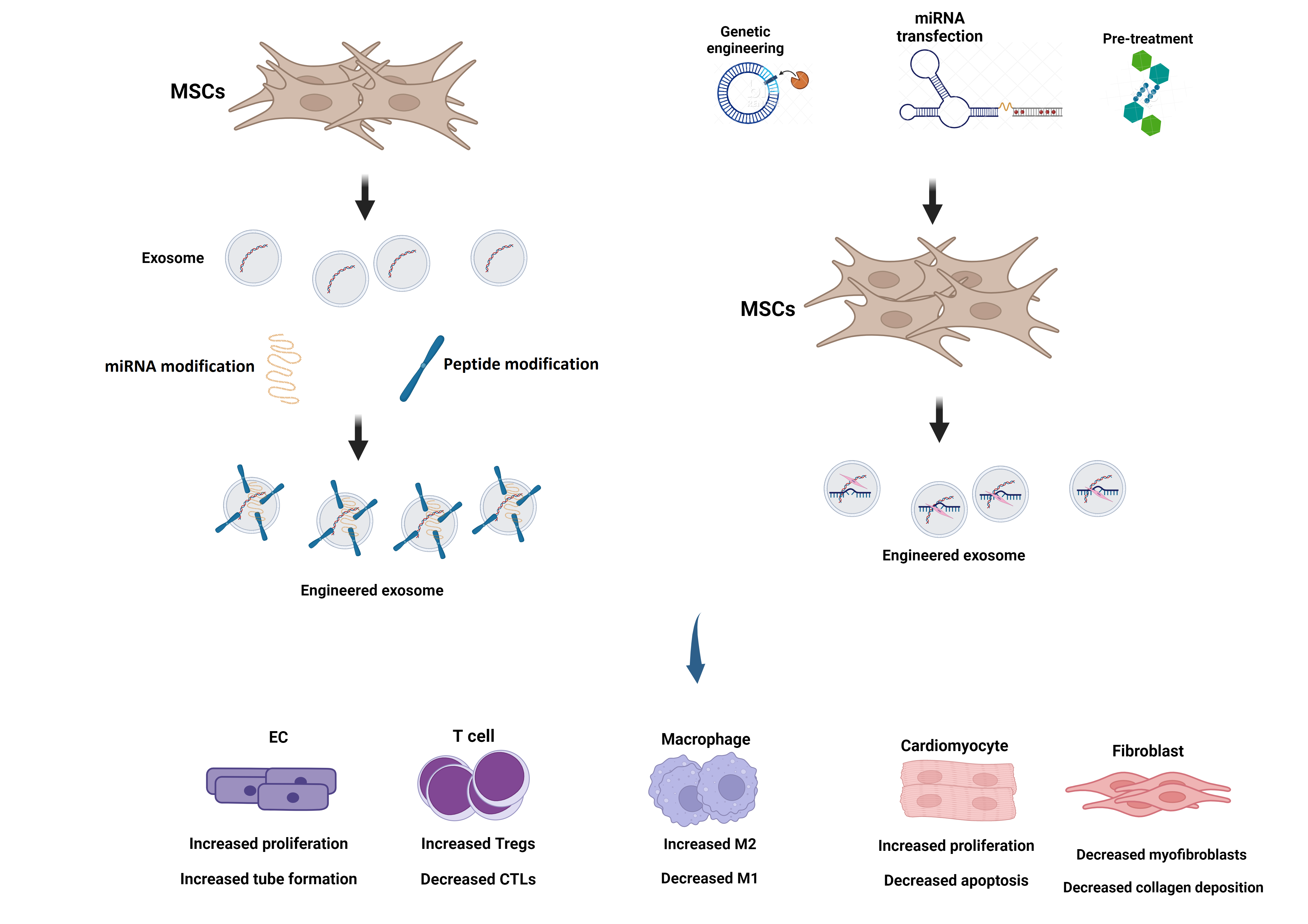
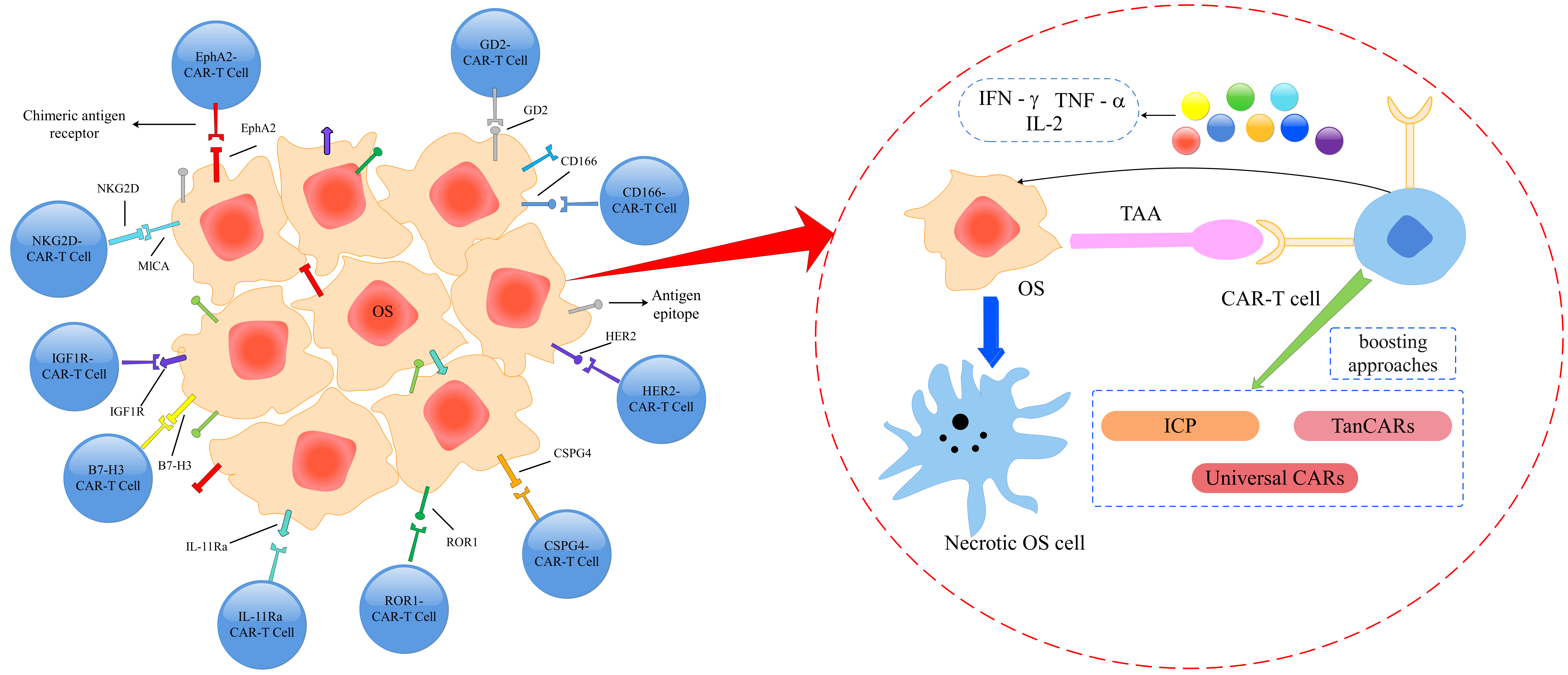
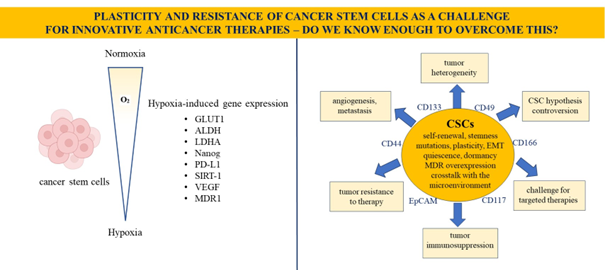
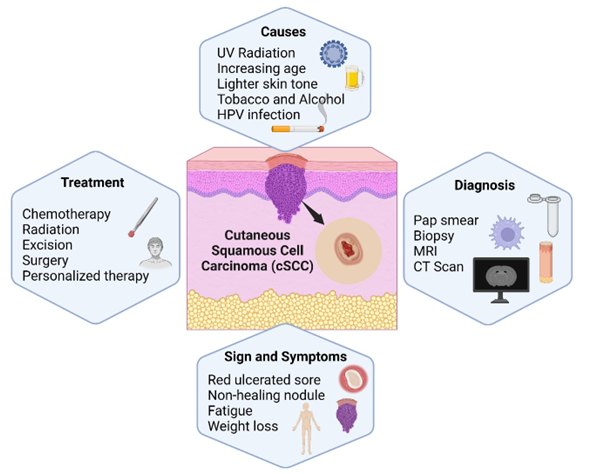
Mandatory Metadata
2024-02-22
Note on submitting a manuscript:
Please check that you have provided all mandatory data!
Read more about Mandatory Metadata
EXCLI Journal publishes original research reports, authoritative reviews and case reports of experimental and clinical sciences.
EXCLI Journal (eISSN 1611-2156) is particularly keen to keep a broad view of science and technology, and therefore welcomes papers which bridge disciplines and may not suit the narrow specialism of other journals. Although the general emphasis is on biological sciences, studies from the following fields are explicitly encouraged:
Immunology, toxicology, ergonomics, neurosciences, psychology, occupational medicine, clinical and preclinical studies, drug development, pharmacology, environmental health, chemistry including analytical chemistry, biochemistry, cell biology, genetics, forensic medicine, oncology and cancer research, proteomics, systems biology, hepatology and gastroenterology, aging research, psychiatric research, behavioral sciences.
Note on submitting a manuscript:
Please check that you have provided all mandatory data!
Read more about Mandatory MetadataPublished continuously
Categories (from 2022 onwards)
Cancer Research Human Factors Immunology Life Sciences Neuroscience Pharmacology/Toxicology
Powered by
![]()

EXCLI Journal (eISSN 1611-2156) is a platinum open access journal. There are neither fees for authors submitting their papers nor fees for readers accessing PDFs of the published papers.
Articles published in EXCLI Journal are licensed under a Creative Commons Attribution 4.0 International License.