Research article
Evaluation of glutathione S-transferase P1 (GSTP1) Ile105Val polymorphism and susceptibility to type 2 diabetes mellitus, a meta-analysis
Mostafa Saadat1[*]
1Department of Biology, College of Sciences, Shiraz University, Shiraz 71467-13565, IranEXCLI J 2017;16:Doc1188
Abstract
It is well established that type 2 diabetes mellitus (T2DM) is associated with oxidative stress and glutathione S-transferases (GSTs) protect cells against oxidative stress. The missense substitution Ile105Val (rs1695) of the glutathione S-transferase P1 (GSTP1, OMIM: 134660) results from an A/G base substitution at nucleotide 313. Many studies have evaluated the correlation between the rs1695 polymorphism and T2DM, but the results remain inconclusive. The aim of the present meta-analysis was to investigate the association between GSTP1 Ile105Val polymorphism and the susceptibility risk of T2DM. Eligible studies (published before August 2017) were identified in several databases. The heterogeneity between studies was evaluated with the chi-square based Q test and the I2 test. The strengths of the association were assessed by pooled odds ratios (ORs) and the corresponding 95 % confidence interval (95 % CI) using either a fixed or random-effects models. Eighteen studies documenting a total of 2595 T2DM cases and 2888 controls were included in this meta-analysis. In the overall analysis there was no significant association between the rs1695 polymorphism and the risk of T2DM. The subgroup analyses stratified by ethnicity, publication year and sample size did not reveal significant association between the study polymorphism and the risk of T2DM and any sources contributing to the substantial heterogeneity between studies. The present meta-analysis suggested that there was significant heterogeneity between studies. Considering some limi tations of our meta-analysis, further large-scale studies should be done to reach a more comprehensive understanding.
Keywords: susceptibility, heterogeneity, risk, Diabetes mellitus, genetic polymorphism
Introduction
Oxidative stress defines as imbalance between reactive oxygen species and anti-oxidant defense systems and it is associated with pathogenesis of diseases. In human, genes involved in antioxidant defenses are highly polymorphic and show association with several multifactorial traits (Saadat, 2006[25], 2013[26]; Saadat and Saadat, 2015[27]; Kang, 2015[15]; Jamhiri et al., 2017[13]; Zendehboodi, 2017[37]).
It is well established that oxidative stress plays an important role in the development of type 2 diabetes mellitus (T2DM) (Asmat et al., 2016[5]). The prevalence of T2DM is rising rapidly in both developed and developing countries (Ginter and Simko, 2012[9]; Sun et al., 2014[30]). Many studies point to that genetic predisposition plays an important role in the development of T2DM (Almgren et al., 2011[3]; Sun et al., 2014[30]; Stančáková and Laakso, 2016[28]).
One of the most important cellular detoxification systems is glutathione S-transferase (GST, EC 2.5.1.18) enzymes. The GSTs are involved in catalyzing the conjugation reactions of reactive intermediates of electrophilic compounds with cytosolic glutathione. The glutathione S-transferase P1 (GSTP1, OMIM: 134660) encodes the π-class of GSTs, which accounts for about 90 % of the enzymatic activity of the GST family; and expressed in many normal tissues (Townsend and Tew, 2003[32]).
Human GSTP1 is polymorphic and its variant alleles occurr at a high frequency. The missense substitution Ile105Val (rs1695) of the GSTP1 results from an A/G base substitution at nucleotide 313. The Val105 form of the GSTP1 enzyme may be 2-3 times less stable than the Ile105 form (Johansson et al., 1998[14]) and may be associated with a higher level of DNA adducts (Ryberg et al., 1997[24]). Enzymes with the valine at amino acid 105 have a seven-fold higher catalytic efficiency for the diol epoxides of polycyclic aromatic hydrocarbons than the isoenzymes with the isoleucine at this position. In contrast, the Val105 enzyme is three-fold less effective with 1-chloro-2,4-dinitrobenzene as a substrate (Zimniak et al., 1994[38]; Hu et al., 1997[12]; Sundberg et al., 1998[31]).
The association between the Ile105Val GSTP1 polymorphism and the risk of T2DM has been investigated, but these studies yielded controversial results (Yalin et al., 2007[35]; Oniki et al., 2008[20]; Bid et al., 2010[6]; Tsai et al., 2011[33]; Ramprasath et al., 2011[21]; Amer et al., 2012[4]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]). Some studies suggested that the GSTP1 polymorphism is associated with susceptibility to T2DM (Bid et al., 2010[6]; Amer et al., 2012[4]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]), other reports, however, do not support the finding (Yalin et al., 2007[35]; Oniki et al., 2008[20]; Tsai et al., 2011[33]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Abbasi et al., 2014[1]; Rasheed et al., 2016[23]). Whether GSTP1 polymorphism modifies the risk of T2DM remains uncertain, therefore the present meta-analysis was carried out.
Methods
Search strategy
Literature databases, including PubMed, Scopus, DOAJ, Index Copernicus, JSTOR, JournalTOCs, AJOL (African Journal Online), Asia Journal Online, Index Scholar, Academic Journals Databases, Google Scholar, Research Bib, High-Wire, J-STAGE, Serbian citation index, KoreaMed, IndMed, PakMediNet and SID (Scientific Information Database) were searched for relevant studies (the last search was updated in August 2017).
The following search terms were used: ('type 2 diabetes mellitus', 'glutathione S-transferase P1' 'GSTP1', 'rs1695', polymorphism, Ile105Val. The search was limited to studies published in English or Persian. In addition the bibliographies of the retrieved studies were screened to identify relevant publications.
Inclusion and exclusion criteria
The eligible studies had to meet the following criteria: (1) a case-control study to evaluate the association between rs1695 polymorphism and risk of T2DM; (2) raw data including sample size, allele or genotype distribution were given. Accordingly, the exclusion criteria were as follows: reviews, meta-analysis, editorial articles, abstracts, comments.
Data extraction
The following information was extracted from each: Name of the first author, year of publication, country of origin, ethnicities of the individuals involved, number of cases and controls, genotyping method, and number of genotypes for the study polymorphism in cases and controls.
Statistical analysis
The crude odds ratios (ORs) and 95 % confidence intervals (95 % CIs) of GSTP1 Ile105Val polymorphism and risk of T2DM were estimated for each study. Hardy-Weinberg equilibrium in controls was tested by a chi-square test.
The heterogeneity between studies was evaluated with the chi-square based Q test and the I2 test. An I2 value of less than 25 % indicates low heterogeneity, 25 % to 50 % indicates moderate heterogeneity, and greater than 50 % indicates high heterogeneity. I2> 50 % or P < 0.10 for Q statistics was considered as significant heterogeneity.
If no significant heterogeneity was found between the studies, the pooled OR was calculated by using the fixed effects model (the Mantel-Haenszel method) (Mantel and Haenszel, 1959[16]). Otherwise, the random effects model (the DerSimonian and Laird method) was applied (DerSimonian and Laird, 1986[7]). Three genetic models were performed in the meta-analysis: codominant model (Ile/Val vs Ile/Ile and Val/Val vs Ile/Ile), dominant model (Ile/Val + Val/Val vs Ile/Ile) and allele model (Val vs Ile). We also performed subgroup analysis according to ethnicity (Caucasian), sample size (≤300/>300 subjects), and publication year (prior to or during 2012/after 2012).
Moreover, sensitivity analysis was performed to evaluate the stability of the results following sequential removal of each study. To determine the evidence of publication bias, the funnel plot and Egger's test were both used. An asymmetric plot suggested possible publication bias. For the interpretation of Egger's test, statistical significance was defined as P<0.05 (Egger et al., 1997[8]).
Results
In total, 18 independent studies including 5483 subjects (2595 T2DM cases and 2888 controls) were included in the study. The studies were published between 2007 and 2017. The characteristics of each study were summarized in Table 1(Tab. 1) (References in Table 1: Yalin et al., 2007[35]; Oniki et al., 2008[20]; Bid et al., 2010[6]; Tsai et al., 2011[33]; Ramprasath et al., 2011[21]; Amer et al., 2011[4]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]). The studies included in the present meta-analysis were conducted on different ethnic populations: 14 studies included a Caucasian population, 2 studies included an Asian population, and 2 studies included an African population. In all studies the study polymorphism was evaluated by RFLP-PCR method.
In overall there was no significant association between the study polymorphism and the risk of T2DM under codominant (Figure 1(Fig. 1); References in Figure 1: Yalin et al., 2007[35]; Oniki et al., 2008[20]; Bid et al., 2010[6]; Tsai et al., 2011[33]; Ramprasath et al., 2011[21]; Amer et al., 2011[4]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]), dominant (Figure 2A(Fig. 2); References in Figure 2: Yalin et al., 2007[35]; Oniki et al., 2008[20]; Bid et al., 2010[6]; Tsai et al., 2011[33]; Ramprasath et al., 2011[21]; Amer et al., 2011[4]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]) and allele genetic models (Figure 2B(Fig. 2)). It should be noted that there was significant heterogeneity in the examined genetic models for the rs1695 polymorphism (Table 2(Tab. 2); References in Table 2: Oniki et al., 2008[20]; Vats et al., 2013[34]; Rao et al., 2014[22]; Ahmed and Al-Bachary, 2017[2]). The genotyping frequencies in the control groups showed significant differences with the expected frequencies in three studies (Vats et al., 2013[34]; Rao et al., 2014[22]; Ahmed and Al-Bachary, 2017[2]). After excluding these studies from meta-analysis, no significant association between the Ile105Val polymorphism and the T2DM risk under examined models was observed (Table 2(Tab. 2)). Excluding 3 studies did not alter heterogeneity between studies.
The source of heterogeneity was assessed by ethnicity, publication year, and sample size. The subgroup analyses did not reveal any sources contributing to the substantial heterogeneity (Table 2(Tab. 2)). In stratified subgroups based on ethnicity, publication year and sample size, no statistically significant association was observed between the GSTP1 Ile105Val polymorphism and the risk of T2DM in any of the genetic models (Table 2(Tab. 2)).
Sensitivity analyses were performed by sequential omission of individual studies for all subjects and subgroups. The corresponding pooled ORs were not altered in all subjects and subgroups of GSTP1 genotypes (data not shown). The results of sensitivity analyses indicated the stability of the results of this meta-analysis. Funnel plots were created and Egger's test was performed to assess the publication bias of the included studies. The funnel plots did not show obvious asymmetry in the overall population (data not shown).
Discussion
It has been well established that oxidative stress is involved in pathogenesis of T2DM (Sun et al., 2014[30]). Considering that GSTP1 belongs to GST superfamily and is involved in cellular detoxification and the fact that Ile105Val polymorphism may alter the GSTP1 enzyme activity (Zimniak et al., 1994[38]; Johansson et al., 1998[14]; Ryberg et al., 1997[24]; Hu et al., 1997[12]; Sundberg et al., 1998[31]), it seems that the rs1695 polymorphism may be associated with susceptibility to T2DM. During 2007 to 2017, eighteen studies investigate the association between this polymorphism and the risk of T2DM (Yalin et al., 2007[35]; Oniki et al., 2008[20]; Bid et al., 2010[6]; Tsai et al., 2011[33]; Ramprasath et al., 2011[21]; Amer et al., 2012[4]; Moasser et al., 2012[19]; Gönül et al., 2012[10]; Grubisa et al., 2013[11]; Mastana et al., 2013[17]; Vats et al., 2013[34]; Rao et al., 2014[22]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]). However, the results of the studies are not consistent. Therefore we carried out a meta-analysis. In the present meta-analysis we found high level of heterogeneity between studies and no association between the examined genetic models and the risk of T2DM (Table 2(Tab. 2)).
Some limitations of the present meta-analysis should be acknowledged. The non-conformity of the geographical distribution of the association studies used in the meta-analysis is the most important limitation of the present study. There was no report from America continent, Western Europe, Australia and East of Asia. Second, the sample size in some subgroup analysis was small (Yalin et al., 2007[35]; Tsai et al., 2011[33]; Grubisa et al., 2013[11]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]), which may increase the risk of false negatives or false positives. Third, there was no data regarding the source of control groups in studies used for meta-analysis (Moasser et al., 2012[19]; Grubisa et al., 2013[11]; Abbasi et al., 2014[1]; Zaki et al., 2015[36]; Stoian et al., 2015[29]; Mergani et al., 2016[18]; Rasheed et al., 2016[23]; Ahmed and Al-Bachary, 2017[2]). Therefore we failed to found source of heterogeneity. Fourth, there was still heterogeneity in the subgroup analysis. The more confounding factors should be considering. Finally, numerous environmental, genetic factors and the interactions among these factors contribute to the progression of T2DM. Our meta-analysis results did not adjust any confounding covariant, for example gender, drinking status, smoking habit.
A more comprehensive analysis should be conducted when more original information is available and interactions among the risk factors are considered. Further well-designed large studies are required to investigate gene-environment interactions.
Acknowledgement
This study was supported by Shiraz University.
Conflict of interest
No competing interests are declared by any of the authors.
References
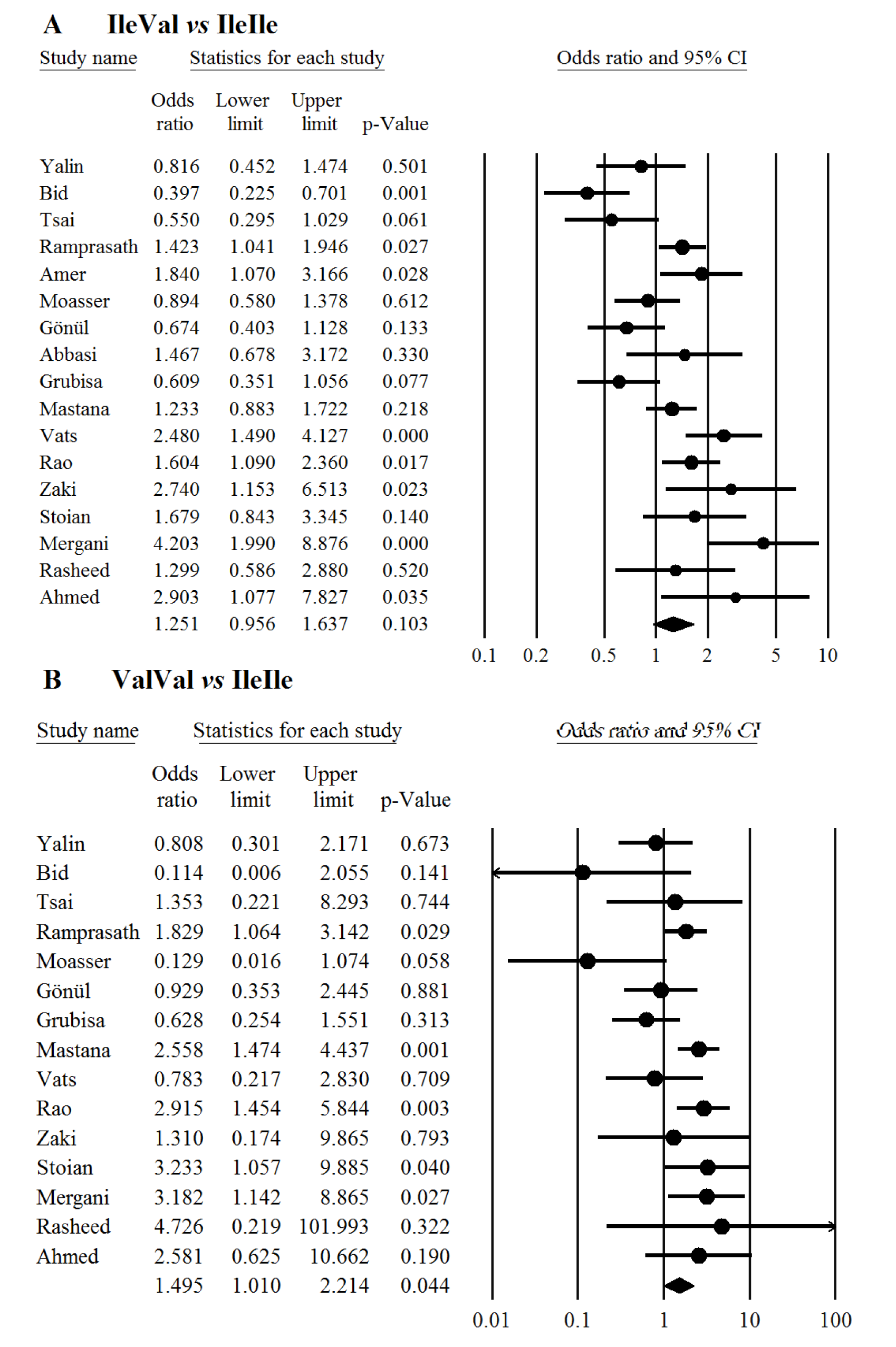
Figure 1: Meta-analysis of the GSTP1 Ile105Val polymorphism with type 2 diabetes mellitus under codominant genetic model; Panels A and B represent Ile/Val vs Ile/Ile and Val/Val vs Ile/Ile respectively. Each box represents the odds ratio (OR) point estimate and its 95 % confidence interval (CI). The diamond represents the overall summary estimate, with confidence interval represented by its width. Pooled ORs and 95 % CI are estimated by Dersimonian and Laird random-effects model.
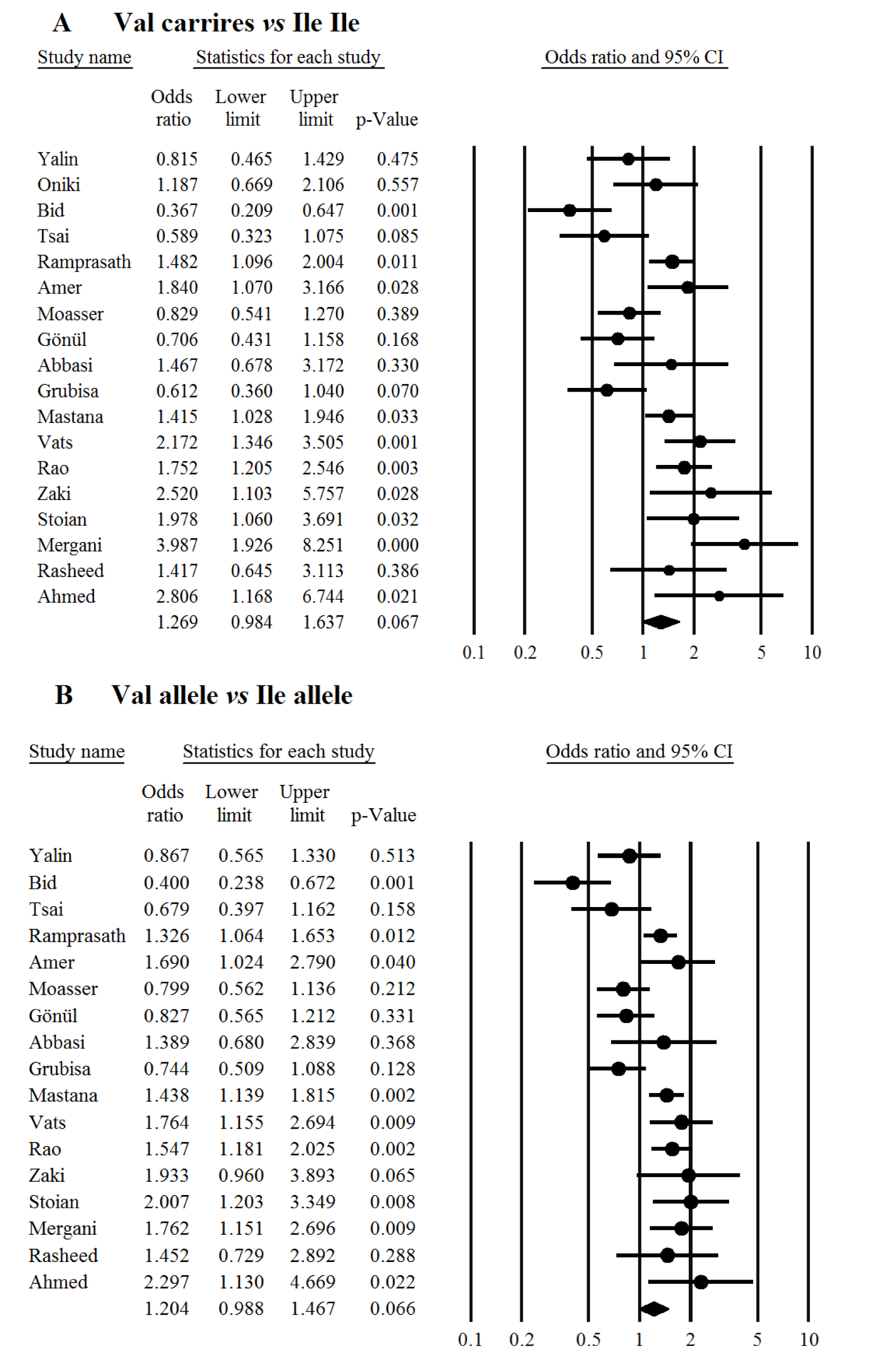
Figure 2: Meta-analysis of the GSTP1 Ile105Val polymorphism with type 2 diabetes mellitus under dominant (A) and allele genetic models (B). Each box represents the odds ratio (OR) point estimate and its 95 % confidence interval (CI). The diamond represents the overall summary estimate, with confidence interval represented by its width. Pooled ORs and 95 % CI are estimated by Dersimonian and Laird random-effects model.
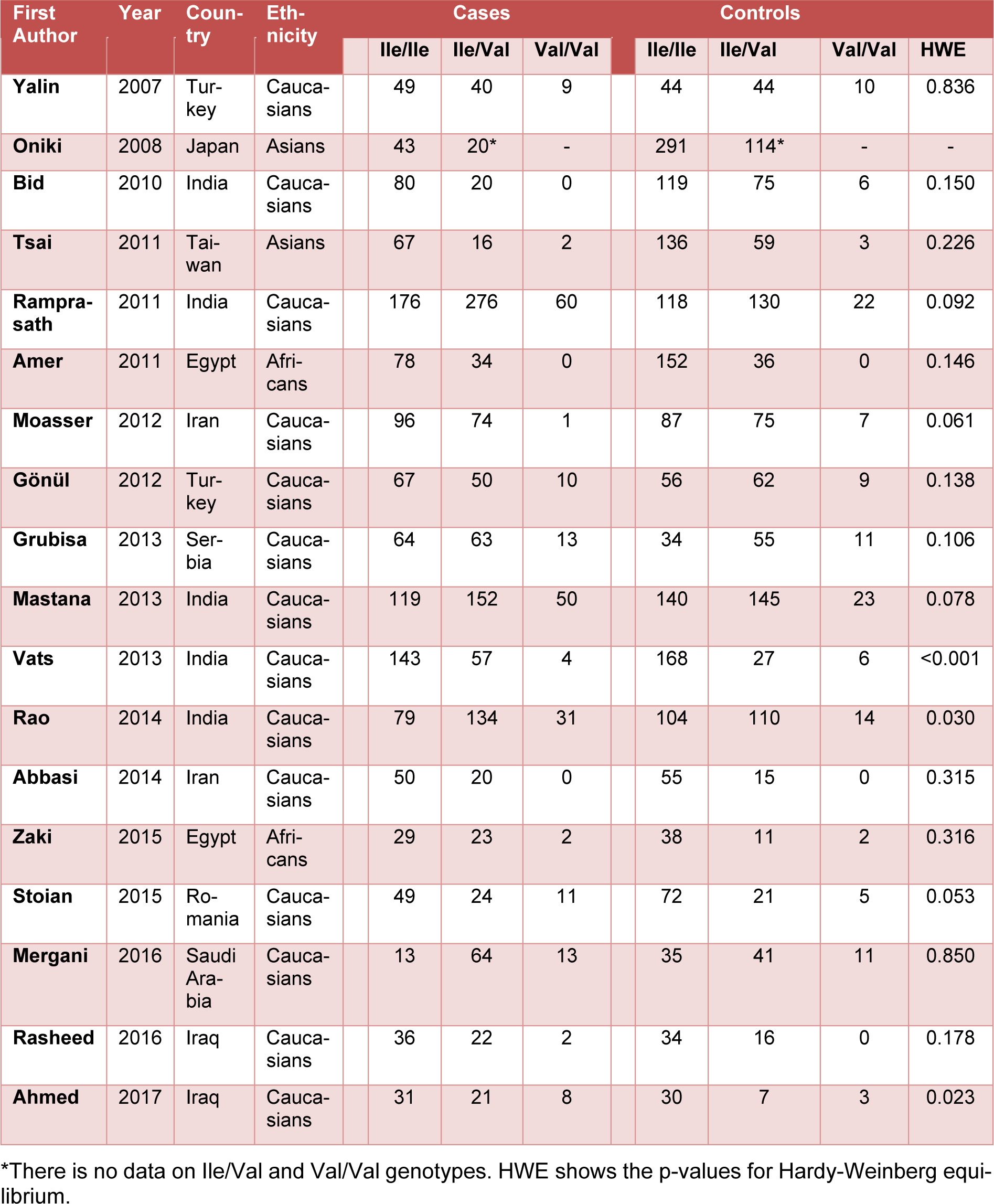
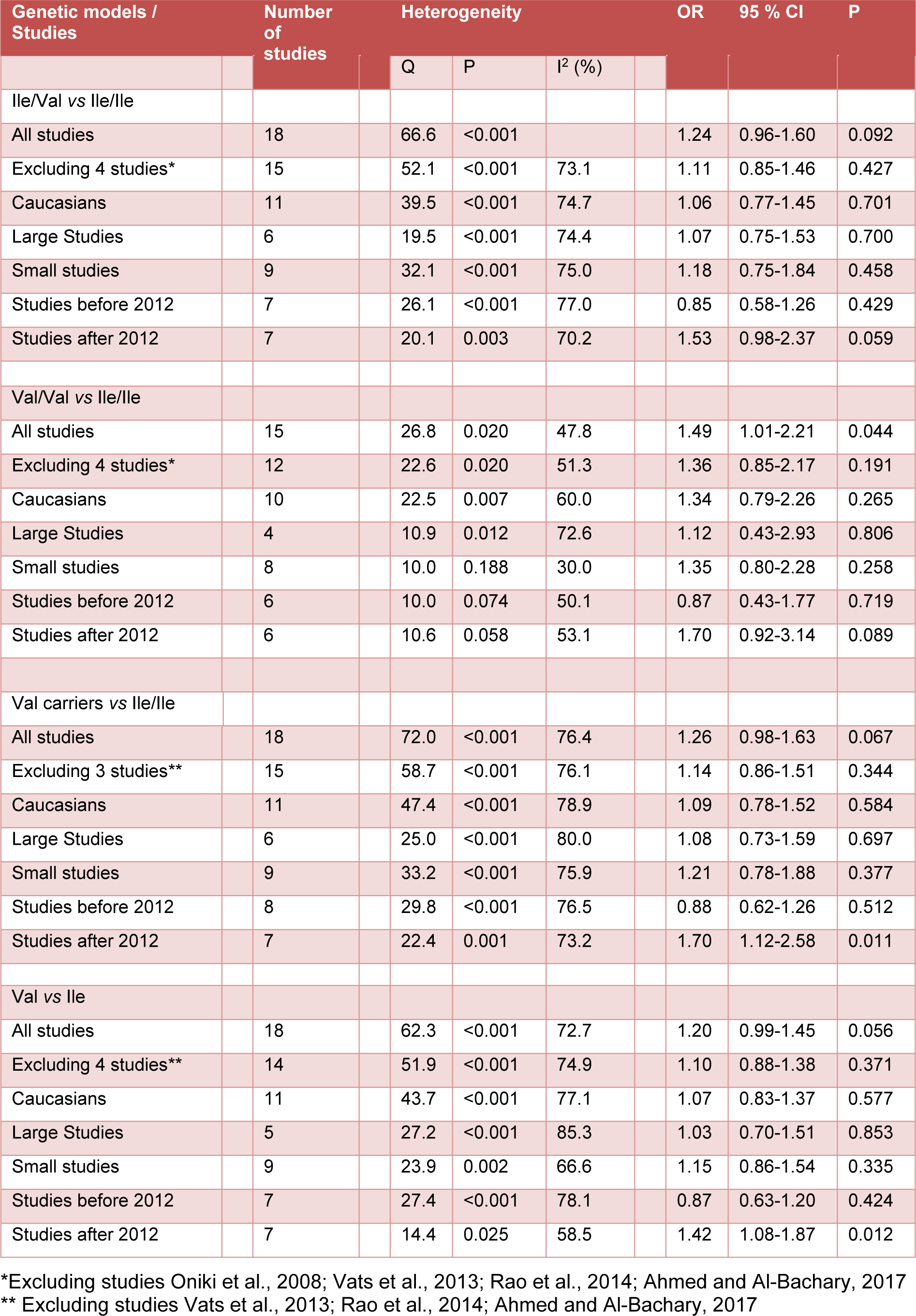
Table 2: Summary of meta-analysis of studies evaluating GSTP1 Ile105Val polymorphism and type 2 diabetes mellitus
[*] Corresponding Author:
Mostafa Saadat, Department of Biology, College of Sciences, Shiraz University, Shiraz 71467-13565, Iran; Tel: +98-71-36137432, Fax: +98-71-32280916, eMail: saadat@shirazu.ac.ir and msaadat41@yahoo.com